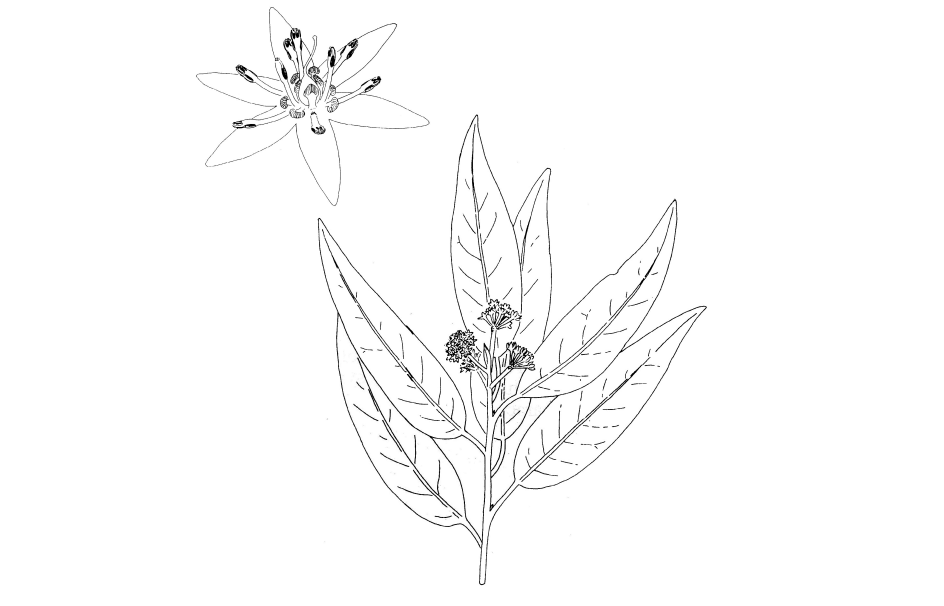
California Bay Laurel
Bonnie’s cover drawing this time is a modified repeat from May 2009. It is derived from one that she did for David Keil and my plant taxonomy text. My guess is that it is a tree that almost all of you know already. It is one of the first trees for which I learned its name. It is known locally as the California bay laurel or simply California bay. Its scientific name is Umbellularia californica and belongs to the laurel, sassafras, cinnamon or camphor family (Lauraceae). As can be surmised from the drawing of a flowering twig tip, it produces small flowers. Each yellowish-green flower cluster turns into a single dry olive-like fruit.
Why discuss this species so soon?
It’s because Heather asked me to explain the new family placements in the new Jepson Manual. Up until the middle of the 20th century, the flowering plants were divided into only two taxonomic classes. These were the monocots and the dicots. Different taxonomists divided the flowering plants in various ways, but none seriously messed with the dicot/monocot distinction. Then in the late 1960’s, Arthur Cronquist came up with a new classification for the dicots which accounts for 2/3 of the flowering plants. It should be noted that he too didn’t mess seriously with the two classes – dicot and monocot. What he did do was recognize an evolutionary basal subclass he called the Magnoliidae.
This subclass contained many woody plants which displayed characteristics that he considered very primitive. These included such traits as a wood anatomy more like conifers than the rest of the flowering plants. A few of them, but none of our native California plants, even had immature seeds (ovules) that were exposed to the open via an opening in their ovaries which resulted in a pollination process where the pollen landed directly on the ovule. Again this is reminiscent of what occurs in gymnosperms. One thing we need to remember about the plants classified in this subclass is that they all produced true flowers so there was no controversy about their being flowering plants.
DNA Sequencing
We now skip ahead to the 1980s and 1990s. Genetic procedures were developed that allowed the molecule DNA (deoxyribonucleic acid) to be readily extracted from organisms and duplicated rapidly. This produced sufficient quantities to be easily studied. Studying DNA means determining the sequences of the four nucleotides that are found in all DNA molecules. These nucleotides include A (adenine), C (cytosine), G (guanine), and T (thymine). Basically all DNA molecules contain long sequences of these four nucleotides in patterns unique to the group to which an individual organism belongs. Each individual within a group also possesses DNA sequences that are a very slight variation of its group DNA.
At this time plant taxonomists combined the newer DNA sequences with older morphological (form or appearance) and biochemical traits (as well as fossil evidence where available) into extremely large data tables (similar to computer spreadsheets and tables produced in Microsoft WORD and EXCEL only larger and read by different software. These huge data sets required computers running specialized analysis programs. These programs basically create groups of species on the basis of similarity using all the characteristics including DNA sequence data. That is, it would first link species together that shared the most characteristics. Then it would combine these new groups, again based on combined similarity, into a smaller number of slightly larger groups.
If you repeat this procedure long enough, it will produce series of fewer but larger clusters. Ultimately, the large number of individual starting groups (species) will end up in a single, all encompassing group. The computer can also produce a picture of the process. This diagram resembles an intricately branched shrub or tree. In the diagram (right) , species (or genera or families) are represented by letters and the numbers represent degree of similarity or percent of shared characters. In the diagram below, the many first-formed, highly similar small groups appear to the right of the tree or at the branch tips while the few last formed, diverse, composite groups appear toward the left. I’m guessing that some of you will picture the tree as potentially representing an evolutionary sequence, with the more primitive groups at the left and the derived (advanced) groups to the left.
, species (or genera or families) are represented by letters and the numbers represent degree of similarity or percent of shared characters. In the diagram below, the many first-formed, highly similar small groups appear to the right of the tree or at the branch tips while the few last formed, diverse, composite groups appear toward the left. I’m guessing that some of you will picture the tree as potentially representing an evolutionary sequence, with the more primitive groups at the left and the derived (advanced) groups to the left.
How does all this impinge on the placement of California bay, as well as spicebush (Calycanthus) and yerba manza (Anemopsis), in Jepson? Well, when this process was repeated many times by many researchers, it turned out that these plants fell not only below and separate from the rest of the dicots and monocots but also between the dicots and monocots. The only way to translate these relationships into a classification system was to create a new category of flowering plants that is neither monocot nor dicot but equal to them in rank. This is the “magnoliids.”
Examine the Bay Laurel
Look at Bonnie’s picture of the enlarged flower. Count the sepals (it doesn’t have any petals). There are six, which is a monocot character. There are also nine stamens also on the monocot 3-merous plan. Note that the plant is a tree whose trunk increases in two diameter via a cylindrical layer of dividing cells (cambium). This, the pinnate veined leaves and two seed leaves (cotyledons) found in the embryo are dicot characters. So even without the esoteric DNA information, a case can be made for the creation of this NEW class of flowering plants to contain intermediates such as our California bay.

Thank you! Please be sure to sign up for email updates. -Judi